
そのため以前には、小さい染色体(B)しかない場合は遺伝子不足によって死亡し、大きい染色体(A)しかない場合は遺伝子過多によって死亡すると考えられていました。
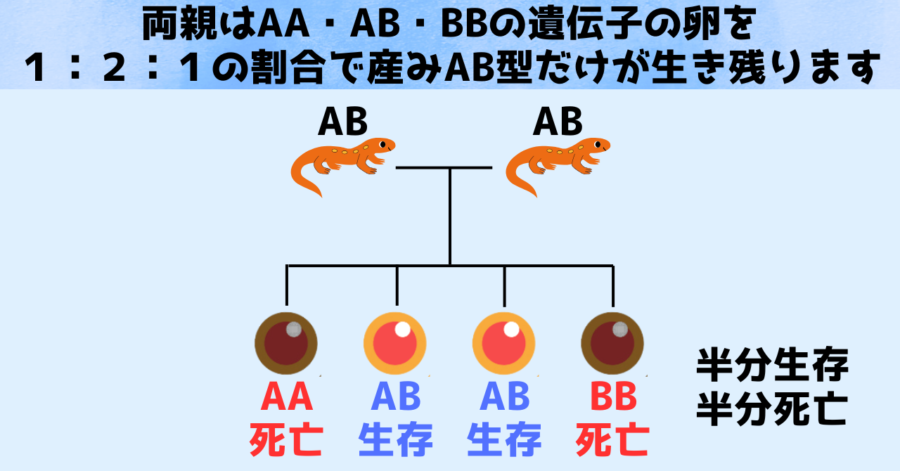
上の図では大きい染色体(A)を2本持つAA型と小さい染色体(B)を2本持つ卵が死に、大きい染色体(A)と小さい染色体(B)を1本ずつ持つ卵だけが生き残る様子を描いています。
中学生で習うメンデルの遺伝法則を第1染色体のみで再現した状態と言えるでしょう。
(※ショウジョウバエなどの研究を行っている研究室では、この致死システムを人工的に再現することで、有害な突然変異の系統を安定的に維持するために使用されています)
また驚くべきことに、この仕組みは均衡致死システムとして知られ、イモリ属の1つTriturus属の9種類全てにみられる特徴となっています。
さらに同じような致死システムは、イモリの異なる種で独自に獲得された可能性も示されています。
本当に不利なシステムならば、種の分岐が起きる前に排除されたり、たまたま採用された1回だけのレアケースになってもよさそうなものですが、そうではなかったのです。
これまでの有力な説では、大きな染色体(A)と小さな染色体(B)はもともとは系統の異なる性染色体であり、致死的なシステムは段階を踏むようにして徐々に形成されていったとされていました。
卵が半分死ぬような理不尽なシステムが急に出現しても、あっという間に絶滅してしまうため、変化は少しずつ進行していったとする説です。
また系統の異なる性染色体であれば、そもそもの大きさが異なっていても説明がつきます。
ただこの説を証明するには、染色体に対する詳細な分析が必要です。
そこで今回ライデン大学の研究者たちは、致死システムを採用するイモリの染色体に対して4226個ものDNAマーカーを打ち込み、詳細な遺伝子地図を構築しました。









































