
また同時に致死システムを採用していないイモリの染色体に対しても同様の遺伝子地図を構築し先祖の代理として扱います。
そして両者の遺伝子地図の比較を行いました。
すると致死システムの獲得は徐々に行われたのではなく、1度の突然変異がキッカケになっていることが判明します。
この結果は、既存の有力な説に反するものです。
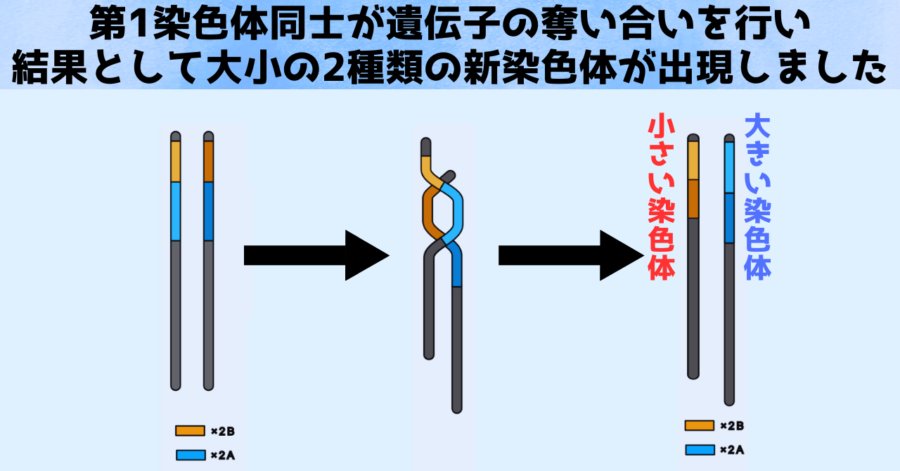
さらに第1染色体に対する分析を行ったところ、大きな染色体(A)と小さな染色体(B)はお互いの遺伝子の奪い合いを行っていたことも判明します。
上の図はその過程を示したものになります。
大きな染色体(A)と小さな染色体(B)はもともと同じ先祖型でしたが、遺伝子を奪い合った結果、大きな染色体(A)はA連鎖遺伝子(寒色)と呼ばれる領域を獲得し、小さな染色体(B)はB連鎖遺伝子(暖色)と呼ばれる領域を獲得しました。
一方の染色体が他方より大きいのは、この奪い合いが同等の割合ではなく不均衡であったことが原因だったのです。
さらにこれらの連鎖遺伝子たちは、相手に存在した自分と同じ遺伝子を多数含んでいることが判明しました。
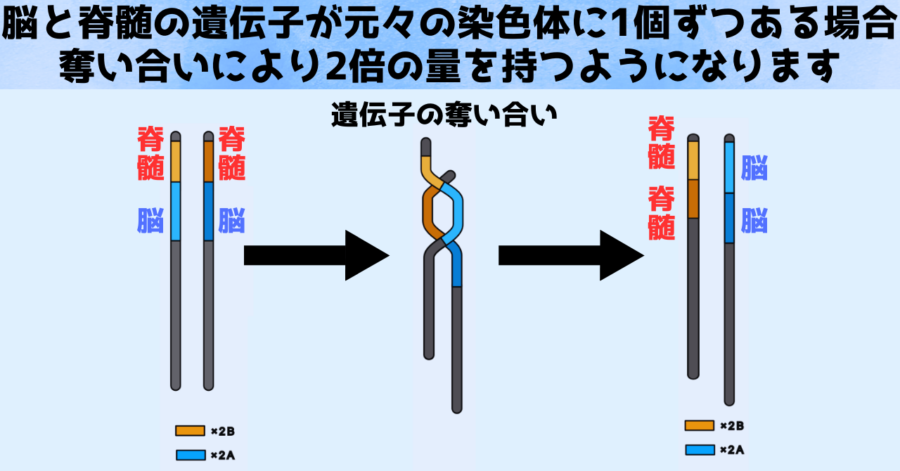
たとえば父親由来の染色体と母親由来の染色体の両方に、脳にとって重要な大きな遺伝子と脊髄にとって重要な小さな遺伝子が1つずつ含まれていたとします。
ですが染色体同士の間で奪い合いが起こった結果、大きな染色体(A)は脊髄の遺伝子を失う代わりに脳の遺伝子を2重に獲得し、小さな遺伝子(B)は脳の遺伝子を失う代わりに脊髄の遺伝子を2重に獲得したとします。









































